
·
Es asincrónica,
Holoblástica, Simétrica, Rotacional(divisiones meridionales y ecuatoriales)
·
A lo largo de este tiempo,
el embrión todavía rodeado por la zona pelúcida, es transportado por la trompa
de Falopio y llega al útero, 6 días después se desprende de la zona pelucida y
se adhiere al revestimiento uterino.
·
Las primeras divisiones
tardan hasta 24 horas c/u
·
Las blastómeras son
totipotenciales hasta el estadio de 8 células
·
MÓRULA.- Cuando el cigoto
consta de 16 células
·
Blastocisto.- cuando el
cigoto consta de 52 células

· Tras varias divisiones entran a una fase llamada COMPACTACIÓN
- Las blastómeras más externas se adhieren íntimamente entre si mediante uniones nexo y estrechas, perdiendo su identidad individual cuando se las observa desde la superficie.
- Esta mediada por la concentración de moléculas de adhesión celular como la E- CADHERINA
- Permite definir entre exterior e interior además de una selección de lo que puede entrar a la mórula.
- Las células externas formarán el TROFOBLASTO, tejido especializado que originará la conexión entre el embrión y la madre.
- Las células del interior formaran la MASA CELULAR INTERNA, que formará al embrión

La actividad de un sistema de transporte de Na basado en la ATPasa de Na/K permite que el Na y agua atraviesen las blastómeras internas, se da 4 días después de la fecundación y se forma una cavidad interna o blastocele. A este proceso se le conoce como CAVITACIÓN.
o En esta fase el embrión consta de una capa
epitelial externa TROFOBLASTO, que dará origen a estructuras extraembrionaria,
incluidas las capas de la placenta, que rodea a la MASA
CELULAR INTERNA, que dará originen al cuerpo del embrión y
otras estructuras extraembrionarias
o El extremo del blastocisto que contiene a
esta última se llama POLO EMBRIONARIO
y el extremo opuesto POLO ABEMBRIONARIO.
·
Existen pruebas de que el FACTOR DE CRECIMIENTO FIBROBLÁSTICO-4,
secretado por la masa celular interna participa en el mantenimiento de la
actividad mitótica den el trofoblasto que la cubre.
Biología
y genética molecular
·
La proteína oct-4 derivada de la madre es necesaria
para permitir que prosiga el desarrollo hasta la fase de dos células.
·
Además en el
establecimiento de las células germinales y la conservación de su
pluripotencialidad.
·
Se expresa en todas las
blastómeras hasta la fase de Mórula y a medida de que comienzan a sufrir varios
tipos celulares diferenciados en el embrión, su nivel de expresión disminuye
hasta que deja de ser detectable.
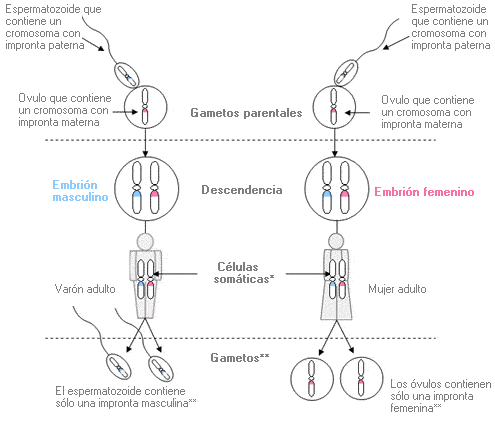
Impronta
Parental
·
Se le denomina así a la
expresión de ciertos genes derivados del óvulo que difieren de la expresión de
los mismos genes cuando derivan del espermatozoide (IGF)
·
Si se retira un pronúcleo
masculino y se reemplaza por otro femenino, el embrión en sí mismo se
desarrolla con normalidad pero la placenta y el saco vitelino hacen lo hace de
forma deficiente
·
Un cigoto con 2 pronúcleos
masculinos origina un embrión con problemas graves de crecimiento mientras que
la placenta y el saco vitelino son casi normales.
·
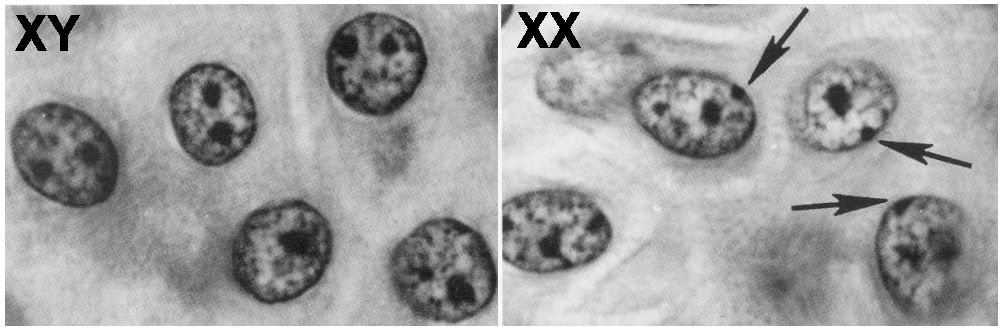
Se sabe que uno de los dos
cromosomas X está inactivado en las células femeninas por su condensación
extrema. Este es origen de la cromatina sexual o CORPÚSCULO DE BARR.
o Los dos componentes del par experimentan una
transcripción activa durante la segmentación temprana de los embriones
femeninos.
o Tras la diferenciación de las blastómeras en
células de trofoblasto o de la masa celular interna, ambos cromosomas continúan
activos en las células de la masa celular interna mientras que en todas las del
trofoblasto el cromosoma X derivado del padre es inactivado de forma selectiva.
o Al final dicha inactivación se produce en
todas las células y sólo durante la ovogénesis se activan de nuevo ambos
cromosomas X
·
Se inicia en el centro de
inactivación de X (CIX), locus exclusivo de este cromosoma.
·
El XIST (transcrito
específico de X inactivo), uno de los genes del CIX, produce un gran ARN sin
capacidad para codificar proteínas, este ARN permanece en el núcleo y cubre el
cromosoma X inactivo por completo, con lo que no permite ninguna trascripción
posterior a este.
·
En el cromosoma X
inactivado el XIST se metila y no se expresa, mientras en el X activo está desmetilado
y transcrpcionalmente activo.
·
El gen XIST es responsable
de la inactivación del cromosoma X paterno en el trofoectodermo y endodermo
extraembrionarios durante la segmentación, mientras que las células de la masa
celular interna no sufren la inactivación del cromosoma X hasta más tarde y
dicha inactivación afecta uno de los cromosomas X al azar.
Propiedades
del desarrollo de los embriones en el periodo de segmentación
·
La REGULACIÓN es la
capacidad de un embrión o del esbozo de un órgano para dar lugar a una
estructura normal cuando se le ha añadido o se han eliminado partes del mismo,
es decir, los destinos de las células de un sistema regulador no están fijados
de forma irreversible y que éstas pueden responder a las influencias
ambientales.
Transporte e Implantación del Embrión
Transporte
por la tuba uterina
·
Toda la etapa inicial de
segmentación tiene lugar mientras el embrión es transportado desde el lugar de
fecundación hasta su sitio de implantación en el útero.
·
La corona radiada se pierde
2 días después de empezar la segmentación, la zona pelúcida se mantiene intacta
hasta que se alcanza el útero.
·
Permanece en la parte
ampular unos 3 días, atraviesa la porción ístmica en 8 horas, gracias a la
progesterona la unión uterotubárica se relaja permitiéndole la entrada en la
cavidad uterina
·
Dos días más tarde (6-7
días después de la fecundación) el embrión se implanta en la porción media de
la pared posterior del putero.
 Zona
Pelúcida
Zona
Pelúcida
·
La disolución de la ZP es justo antes del inicio de
la implantación.
·
Al entrar a la cavidad
uterina la ZP cambia su composición gracias a aportaciones del las blastómeras
y los tejidos reproductores maternos, dichos cambios facilitan el transporte y
la diferenciación del embrión
·
El blastocisto sale gracias
a un orificio provocado por una enzima similar a la tripsina que es secretada
por las células trofoblásticas.
Implantación
en el revestimiento uterino
·
El embrión comienza a
adherirse con firmeza al revestimiento epitelial del endometrio
o Se da gracias a la acción de varias moléculas
de adhesión como las INTEGRINAS.
o Se produce en el área por encima de la masa
celular interna (Polo embrionario).
·
Posteriormente se sumerge
en el estroma endometrial.
o Las células derivadas de este trofoblasto
celular (Citotrofoblasto) se fusionan para formar un sincitiotrofoblasto
multinucleado.
o Las pequeñas prolongaciones del
sincitiotrofoblasto se introducen entre las células epiteliales uterinas,
después se extienden a lo largo de la cara epitelial de la lámina basal que
subyace al epitelio endometrial para formar una placa trofoblástica algo
aplanada.
o El sincitiotrofoblasto inicial es un tejido
sumamente invasivo, que se abre camino con rapidez erosionando el estroma
endometrial, en 10 o 12 días tras la fecundación el embrión está incluido por
completo en el endometrio.
o Mientras el embrión perfora el endometrio y
algunas células citotrofoblásticas se fusionan en el sincitiotrofoblásto, las
células de tipo fibroblástico del estroma endometrial se hinchan por la
acumulación de glucógeno. Estas células deciduales se adhieren de manera muy
apretada y forman una gran matriz celular que primero rodea al embrión
implantado y más tarde ocupa la mayoría del endometrio, a este proceso se le
llama REACCIÓN DECIDUAL.
o Aún no se conocen los mecanismos exactos,
pero al mismo tiempo los leucocitos secretan INTERLEUCINA-2 que parece evitar
el reconocimiento materno del embrión como un cuerpo extraño durante las primeras
etapas de la anidación.
No hay comentarios:
Publicar un comentario